 哥伦比亚大学(Columbia University)和麻省理工学院(MIT)的两个研究小组分别表明,利用CRISPR和转座因子将外源DNA序列插入基因组是可能的。图片由Marco Verch (CC by 2.0) https://www.flickr.com/photos/30478819@N08/47710379171 https://foto.wuestenigel.com/typewriter-blanko-usco-registration/
哥伦比亚大学(Columbia University)和麻省理工学院(MIT)的两个研究小组分别表明,利用CRISPR和转座因子将外源DNA序列插入基因组是可能的。图片由Marco Verch (CC by 2.0) https://www.flickr.com/photos/30478819@N08/47710379171 https://foto.wuestenigel.com/typewriter-blanko-usco-registration/
为了提高CRISPR的效率和准确性,两个研究小组——一个来自哥伦比亚大学,另一个来自麻省理工学院和哈佛大学——各自独立地表明,利用与转座子元件结合的CRISPR酶将DNA序列插入基因组是可能的。这一突破丰富了CRISPR知识库,使其能够以一种有针对性的方式将序列插入染色体。
典型的基因组编辑依赖于DNA修复
改变一个有机体的基因组不是一件容易的事。细胞具有安全机制,旨在纠正由诱变因子(如辐射或化学诱变因子)或在DNA复制过程中错误引入的DNA序列中的错误。DNA修复机制是复杂的酶系统,由环境条件或DNA损伤激活,并保护DNA或试图恢复其原始序列。常见的修复机制有同源重组、非同源端连接、基底切除修复等。这些机制并不完美,允许突变延续到下一代,这对进化过程具有关键意义。
基因组编辑是棘手的,因为它需要考虑、绕过或利用DNA修复系统。例如,酵母和苔藓利用非常高效的同源重组机制,很容易将异种序列合并到它们的基因组中,并执行在活的有机体内基因组组装-当有足够的序列同源性。相反,大肠杆菌通常用于克隆的菌株有其修复蛋白RecA基因失活;这通过减少基因重排提高了插入载体的稳定性。
![]()
举例说明如何使用不同的修复机制进行基因组编辑。非同源端连接(NHEJ)在断裂后连接双链DNA片段。当外源DNA片段存在时,可以合并(A和B)。同源重组(Homologous Recombination, HR)依赖于序列的同源性,可以插入或重新排列基因组中的序列。图像中丹尼尔和曹, 2014, PLOS Biology (CC BY 4.0)
基于crispr的基因组编辑系统需要本地DNA修复系统的合作来完成基因修改。核酸酶,通常是CRISPR Cas9,以非常精确的方式造成DNA损伤。修复酶修复损伤,它们可以通过非同源端连接或同源重组(如下图所示)将异体DNA(需要共渗透到细胞中)合并进来。

使用CRISPR/Cas9进行序列插入。图的Khadotia等, 2016,植物科学前沿(CC BY 4.0)
这种剪切和重新连接在很大程度上导致了CRISPR技术的低效率和突变(另一个重要因素是CRISPR对其他DNA序列的非特异性靶向)。它是可以使用的进行碱基编辑没有减少。然而,该技术依赖于DNA碱基替换,不适合DNA插入或大规模编辑。
在没有双链切割的情况下进行DNA插入或其他修饰可以增强CRISPR技术。两个美国研究小组试图做到这一点,他们将CRISPR和转座因子结合起来。
转座子,又名跳跃基因
转座的元素转座子(也被称为“跳跃基因”)是一种DNA序列,具有移动到基因组不同位置的独特特性。它们最初是由问麦克林托克芭芭拉这一发现使她获得了1983年的诺贝尔生理学或医学奖。
转座子值得获得诺贝尔奖,因为它们对物种的进化做出了贡献。它们会引起基因的复制、缺失和插入,它们可以是水平基因转移的一种手段。它们是遗传学家的宝贵工具,因为转座子库可以帮助确定至关重要的基因或特定遗传要素的遗传适应性和环境适应性.在合成生物学中,转座子实验是构建人工染色体和最小基因组的起点。
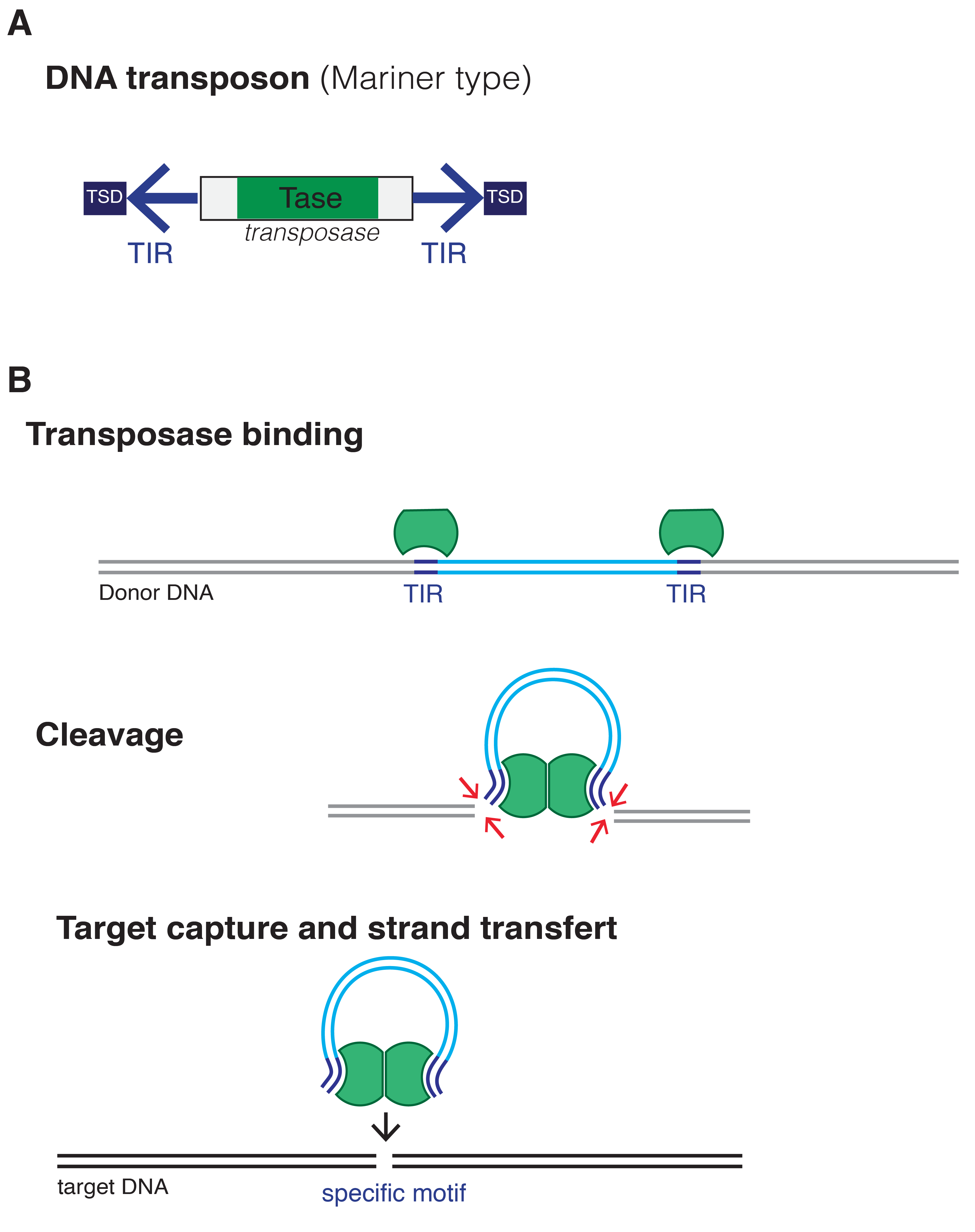
转座子有很多种,它们使用不同的机制进行整合。一个转座子可能使用剪切粘贴的方式,也可能使用逆译,它可能是自主的或非自主的(根据元件是否包含其自身转座子所需的基因),它可能是物种特异性的,也可能在不同的生物体中工作。的Mariner-type转座子下图是在果蝇中首次观察到的果蝇但可以整合到几个不同生物体的基因组中,包括人类。
![]()
II型转位元件的结构及其转位机制。转座酶识别侧翼序列,切除元件,并将其插入一个新的遗传位点。图像Mariuswalter (CC by - sa 4.0)
CRISPR和转座子
2017年,约瑟夫·彼得斯和他的合作者发现一些转座子,特别是像tn7一样的细菌转座子,与它们的细菌宿主的CRISPR系统相关。假设是这些转座元件劫持CRISPR蛋白,以促进其插入基因组。利用这种技术,是否有可能利用CRISPR和工程转座子生成可编程的插入系统?
两个研究小组似乎同意这一观点。来自哥伦比亚大学的Samuel Stenger小组报道这是一项结合了转座子插入和CRISPR精确性的研究。他们称他们的系统
整合(引导rna辅助靶向插入转座元件)。Sanne Klompe和她的合作者从霍乱弧菌, Tn6677。这种跳跃基因利用CRISPR结合对DNA结构造成的干扰,将自己挤进染色体。研究人员描述了该系统的特点,并开发了一个可定制的系统,其中转座子可以插入到不同的遗传位点大肠杆菌。
与此同时,另一组来自麻省理工学院在《科学》杂志报道另一个使用相同原理的插入系统。Jonathan Strecker和他在冯正实验室的合作者在蓝藻中发现了一个紧凑的crispr相关转座子(CAST)Scytonema crispum。研究人员对该系统进行了描述,并对其进行了修改,使其可编程。他们能够将不同长度的DNA序列插入不同的基因座大肠杆菌。报道的效率达到80%,而典型的cas9介导插入的效率仅为1%。

蓝细菌Scytonema crispum(左)和霍乱弧菌(右)(来源:维基共享)
DNA插入问题解决了吗?
这两个研究小组都表明,在理想的染色体位置精确插入DNA序列是可能的,但仅限于细菌。由于CRISPR是一个细菌系统,真核生物中不应该有CRISPR相关的转座元件。整合或CAST系统是否能像在其他生物体中那样发挥作用,或者真核转座子是否能被设计成与非剪切CRISPR蛋白相关联,这将是一件有趣的事情。
有几个基因工程应用可能需要染色体插入长DNA序列,如基因治疗和作物改良。开发一种无需切割的插入系统无疑是迈向更多功能、更安全、更省力CRISPR应用的一步。
6

























{kind=link}
添加评论